EMBRIOLOGIA 1 E 2!
ANEXOS EMBRIONÁRIOS
Os Anexos Embrionários (alantoide, âmnio, cório e vesícula vitelina) são estruturas que se formam a partir dos folhetos embrionários ectoderma, endoderma e mesoderma.
Eles surgem durante a gestação, mas não fazem parte do embrião. Por esse motivo também são chamados de estruturas extra embrionárias e desaparecem com o nascimento.
Têm a função de auxiliar o desenvolvimento do embrião. Isso é feito mediante a cedência de nutrientes, da proteção e da troca entre o embrião e o meio externo através do corpo materno (respiração e excreção).
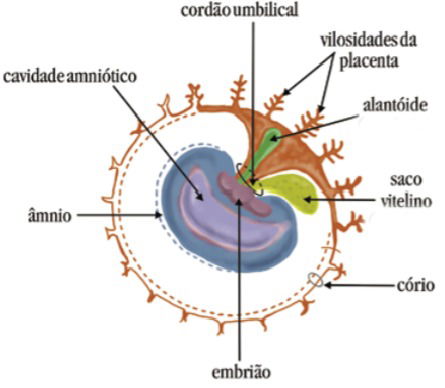
Vesícula Vitelina
A vesícula vitelina, também conhecida como saco vitelínico ou saco vitelino é o primeiro anexo que é formado.
Apresenta aspecto de uma bolsa e surge da endoderma. Além da endoderma, a mesoderma também participa da sua formação, uma vez que a mesoderma reveste a endoderma.
Pelo fato de surgir da endoderma, que é o folheto embrionário que forma alguns órgãos do sistema digestório, a vesícula vitelina está ligada ao intestino do embrião.
Dentro dela está o vitelo, que são os nutrientes que alimentam o embrião. A função da vesícula vitelina é, portanto, nutrir o embrião.
Esse anexo embrionário é muito importante na nutrição das aves, dos peixes e dos répteis. Nos mamíferos, sua função é reduzida, pelo fato de que nesses casos é a placenta que assume esse papel.
Alantoide
O alantoide é uma bolsa que surge da endoderma. Logo, seu exterior é revestido de mesoderma e, tal como a vesícula vitelina, está ligado ao intestino do embrião.
A função do alantoide é armazenar as excretas. As excretas surgem das sobras das substâncias produzidas durante o metabolismo do embrião.
Âmnio
O âmnio tem o aspecto de uma bolsa e envolve todo o embrião. Surge da ectoderma e da mesoderma.
A principal função do âmnio é garantir a hidratação e a proteção do embrião. Além de hidratar, ele absorve o impacto de choques mecânicos e resguarda o embrião para que ele não sofra deformações por aquilo que a Medicina chama de aderência.
Cório
Cório, córion ou serosa é o anexo embrionário que se localiza na parte mais exterior do embrião. É uma membrana que envolve todos os anexos embrionários e que surge da ectoderma e da mesoderma.
Nas aves, pode ser vista numa espécie de pele presente nos ovos.
A função do cório é promover as trocas gasosas, ou seja, garantir a respiração do embrião. Além disso, ele protege o embrião e, no caso dos mamíferos, forma a placenta.
E a Placenta e o Cordão Umbilical
A placenta e o cordão umbilical também são anexos embrionários, mas que estão presentes apenas nos mamíferos.
A placenta é um órgão formado por associação entre tecidos maternos e tecidos embrionários. Ela garante a passagem de nutrientes da mãe para o feto, a troca gasosa e a remoção das excretas.
Tudo isso através do cordão umbilical, que liga a mãe ao feto.
MARSUPIAIS
Os marsupiais (latim científico: Marsupialia) constituem uma infraclasse de mamíferos que se distinguem dos demais de sua classe, como os monotremados e placentários, por sua anatomia e fisiologia reprodutiva: são vivíparos com finalização do desenvolvimento no marsúpio, isto é, são animais que apresentam, em sua maioria e nos indivíduos do sexo feminino, uma dobra de pele a qual origina uma bolsa ou marsúpio provido com glândulas mamárias. Há outras diferenças morfológicas, principalmente reprodutivas, entre elas a presença de duas vaginas na fêmea, e um pênis bifurcado nos machos. Os filhotes nascem precocemente com aparência embrionária e logo se prendem às glândulas mamárias da mãe, a fim de completarem seus desenvolvimentos.
Os marsupiais não são antepassados dos placentários. Ambos os grupos surgiram no Cretáceo e desde então competem pelos mesmos nichos ecológicos. Atualmente, restringem-se à América e Oceania, constituindo cerca de 320 espécies, que correspondem por aproximadamente 6% de todas as espécies de mamíferos. No Brasil, existem 44 espécies, sendo todas pertencentes à família Didelphidae, e 14 gêneros. São conhecidos vulgarmente como "catitas", "cuícas", "Gambás", "guaiquicas" e "marmosas".
Taxonomicamente, o termo Metatheria, proposto por Huxley em 1880, é considerado sinônimo do táxon Marsupialia, proposto por Illiger em 1811 (McKenna e Bell 1997). Entretanto, alguns autores consideram o termo Metatheria mais abrangente, por incluir muitos dos marsupiais primitivos.
CARACTERÍSTICAS
A caixa craniana é pequena e estreita. Abrigando um cérebro relativamente pequeno e simples se comparado àqueles de mamíferos placentários de tamanho similar. O palato é geralmente fenestrado, isto é, contém aberturas em sua superfície óssea. Os marsupiais, caracteristicamente, possuem um ângulo inflectado ao osso dentário, ausente nos eutérios, e seus ossos nasais sobrepõem os ossos frontais com uma forma de diamante, em contraste à forma retangular dos nasais dos eutérios. A abertura do canal lacrimal é ligeiramente anterior à órbita. Usualmente não possuem a bula auditiva, ou podem possuí-la de forma rudimentar, formada a partir de um osso diferente do dos eutérios.
Os marsupiais também diferem dos placentários em sua fórmula dentária. Mesmo entre os marsupiais a fórmula dentária varia consideravelmente, mas basicamente o número de incisivos na maxila superior é diferente do número presente na maxila inferior, exceto na família Vombatidae. O número máximo de incisivos (vistos em várias famílias) é 5/4 em contraste ao 3/3 dos eutérios. O número de pré-molares e dos molares difere também entre os grupos (3/3 e 4/4 nos marsupiais, 4/4 e 3/3 em placentários). O padrão de reposição dos dentes de leite para os permanentes também apresenta ligeira diferença.
O esqueleto pós-craniano dos marsupiais pode ser distinguido dos esqueletos de eutérios, primariamente, por meio dos ossos epipúbicos, os quais são projetados para frente a partir do púbis. Acreditava-se que os ossos epipúbicos eram uma característica única para a sustentação da bolsa (marsúpio); mas se sabe atualmente que eles são um traço primitivo dos mamíferos, retido até mesmo em alguns poucos eutérios primitivos (Novacek et al., 1998). A presença desses ossos epipúbicos é compartilhada com os monotremados.
DISTRIBUIÇÃO E HABITAT
Os marsupiais atualmente são encontrados no continente americano e na região australiana. Entre as Américas, a América do Sul concentra o maior número de espécies, muitas podem também ser vistas na América Central, mas apenas uma espécie ocorre na América do Norte, o gambá-da-Virgínia (Didelphis virginianus). Na região australiana, a maioria das espécies reside na Austrália e Nova Guiné, mas algumas são encontradas nas ilhas Molucas, Sulawesi e ilhas adjacentes.
Os metatérios divergiram da linha dominante dos eutérios no Cretáceo Médio. Acredita-se que as diversificações inicias dos marsupiais ocorreram na América do Norte, entretanto, no período do Mioceno Médio, essa linhagem se extinguiu, somente reaparecendo no período que América do Norte e do Sul foram reconectadas, pela formação do istmo do Panamá, no Plioceno. Alguns poucos fósseis marsupiais são conhecidos da Europa, África e Ásia, mas este grupo nunca foi bem estabelecido nesses continentes.
Os marsupiais estão presentes nos mais variados tipos de habitats, das florestas tropicais úmidas da América do Sul aos desertos australianos.
Os marsupiais exibem vasta formação de comportamentos em consequências de sua evolução para preencher a variedade de nichos ecológicos presentes nos continentes americano e australiano. Sendo assim, uma generalização de seus hábitos se torna difícil. Marsupiais podem ser arbóreos, terrestres, fossoriais e ao menos algumas espécies são semiaquáticas. Quanto aos padrões de locomoção, podem incluir: andar, escalar, cavar, correr ou nadar. Os marsupiais não desenvolveram o processo do voo, entretanto, uma espécie é planadora. Quanto ao padrão de atividades, também existe variedade, podendo ser diurnos, noturnos ou crepusculares. Algumas espécies hibernam, mas muitas permanecem ativas o ano inteiro. Alguns são sociais, enquanto outros são solitários.
Os hábitos alimentares são também sortidos, podendo ser herbívoros, carnívoros, insetívoros, onívoros e nectarívoros.
Marsupiais e placentários diferem fortemente em sua anatomia e padrões reprodutivos. Nas fêmeas, o trato reprodutivo é completamente duplo. Ele consiste de duas vaginas laterais que se unem cranialmente, e a partir deste ponto, os dois úteros separados divergem. As vaginas laterais são apenas para a passagem do esperma. O nascimento dos filhotes se dá por meio de uma estrutura na linha mediana, a vagina mediana ou o canal pseudovaginal, o qual se desenvolve no primeiro parto. Nos machos, o pênis é bifurcado. O escroto, ao contrário dos eutérios, localiza-se à frente do pênis.

Em muitos marsupiais, as fêmeas desenvolvem uma bolsa abdominal, o marsúpio, onde os mamilos estão implantados, tendo também uma função de proteção dos neonatos. O marsúpio não está presente em alguns dasiurídeos (camundongo marsupial) e em alguns didelfídeos (gambás do gênero Didelphis). O marsúpio pode ser vestigial em algumas espécies de tamanho diminuto, como as marmosas.[1]
O período de gestação é muito curto, de 8 a 43 dias dependendo da espécie. Os marsupiais não mantêm o corpo lúteo, e o período gestacional é sempre menor ou igual à duração do ciclo estral, sendo então, os filhotes, ejetados ao final do ciclo éstrico na maioria das espécies. A duração da gestação, nos marsupiais, é relativamente independente do tamanho corpóreo.
O desenvolvimento durante a gestação dos marsupiais é muito diferente do apresentado pelos monotremados e pelos placentários. Os neonatos marsupiais apresentam membros torácicos bem desenvolvidos, seus pulmões são relativamente grandes ao nascimento e diversos aspectos de seu desenvolvimento craniano também são acelerados. O desenvolvimento das mandíbulas, do palato secundário, dos músculos faciais e da língua é avançado (enquanto o do sistema nervoso central é retardado), de forma que um neonato marsupial possa se prender a um mamilo e iniciar a amamentação.

Quando os filhotes nascem, eles devem deixar a vagina para se prender a um mamilo, completando assim seu desenvolvimento. O principal modo observado nos macropodídeos (cangurus) é aquele no qual o neonato escala o corpo da mãe até à bolsa, com a ajuda de seus membros anteriores bem desenvolvidos. Acredita-se que o desenvolvimento desses membros em um estágio tão cedo do crescimento dificulta em sua diferenciação para estruturas como nadadeiras e asas, como é visto em outros grupos de mamíferos[3]. A mãe adota uma postura sentada distinta e lambe o caminho da vagina até a bolsa, mas não ajuda o filhote de nenhuma outra maneira em sua jornada. Alguns dasiurídeos e didelfídeos apresentam filhotes mais altriciais que os cangurus. Estes neonatos são ejetados diretamente para as bolsas (ou então para as áreas mamárias de espécies sem bolsa) no nascimento. Os filhotes recém-nascidos destas espécies são passivos, diferentemente dos cangurus.
A maior parte do desenvolvimento do filhote se dá no marsúpio, sendo assim, o período de lactação excede amplamente o período de gestação. A lactação também continua, por algum tempo, mesmo que o filhote já tenha se desenvolvido o suficiente para se desprender do mamilo da mãe. É nesse momento que tipicamente observamos os filhotes entrando e saindo de suas bolsas. E pode durar até um ano, ou mais em algumas espécies (como no gênero Macropus).
Assim, a fêmea marsupial investe pouca energia e recursos durante a gestação, mas a lactação requer investimento substancial. A composição do leite tem variações marcantes nos marsupiais. O primeiro leite é diluído e rico em proteínas, enquanto o leite posterior é mais concentrado e rico em gorduras. A lactação concomitante sem sincronia foi observada em alguns cangurus (como o canguru-vermelho, Macropus rufus); ou seja, um filhote imaturo preso a um dos mamilos enquanto um filhote mais maduro bebe em outro mamilo, e a mãe produz tipos distintos de leite em cada um dos mamilos. A composição do leite é, provavelmente, determinada pelo tempo que cada mamilo o produz.
Os padrões reprodutivos também variam consideravelmente entre os marsupiais. Muitas espécies são solitárias, encontrando-se somente na época reprodutiva. A poliginia também pode ser observada em alguns grupos, onde machos disputam o acesso a várias fêmeas, como nos coalas, ou então em estruturas hierárquicas com um macho dominante em animais com vida social, como por exemplo o Macropus parryi. A monogamia também está presente, como no Petauroides volans, que vivem em grupos familiares que consistem no casal e suas crias.
Tipos de Óvulos
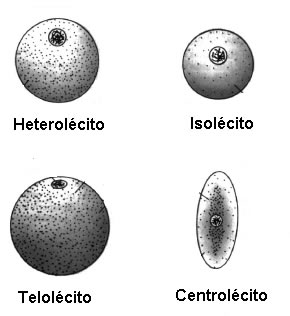
A classificação dos óvulos nos organismos animais varia de acordo com a quantidade e a distribuição de vitelo (substância de reserva energética) presente em seu interior, implicando no padrão de segmentação (ou clivagem) da célula ovo ou zigoto.
Quanto maior a quantidade de vitelo, menor é a velocidade do processo de segmentação;Quanto menor a quantidade de vitelo, maior a velocidade do processo de segmentação.
Isolécito (oligolécito, alécito ou homolécito) → óvulos com baixa quantidade de vitelo, dispersos homogeneamente no citoplasma da célula.
- Tipo de segmentação: holoblástica igual.
- Exemplo: equinidermos, espongiários, protocordados e mamíferos.
Heterolécito (mediolécito ou miolécito) → óvulos com muito vitelo dispersos pela célula, formando uma região distinguindo o pólo animal (com pouco vitelo, contendo o núcleo) e o pólo vegetativo (concentrada em vitelo).
- Tipo de segmentação: holoblástica desigual, formando após divisão, dois tipos de células com tamanhos diferenciados (os micrômeros e os macrômeros).
- Exemplo: moluscos, platelmintos, anelídeos, anfíbios e alguns peixes.
Telolécito (megalécito) → óvulos maiores, contendo muito vitelo disperso na célula. O núcleo fica localizado em uma região delimitada por uma cicatrícula que separa o pólo animal do vegetal.
- Tipo de segmentação: Meroblástica discoidal.
- Exemplo: répteis, aves e alguns peixes.
Centrolécito→ óvulo com vitelo concentrado na região central juntamente ao núcleo. O vitelo nesse tipo de célula não se divide, porém o núcleo passa por diversas divisões com posterior migração para a periferia.
- Tipo de segmentação: Meroblástica superficial.
- Exemplo: artrópodes.
Tipos de segmentação embrionária

Etapas de desenvolvimento do Embrião
MÓRULA
A mórula é o primeiro estágio embrionário. Ela é maciça, repleta de células.
E como ela se forma?
Quando ocorre a fecundação, ou seja, quando o espermatozoide penetra no ovócito, vários eventos acontecem. Um evento importante é a fusão de dois pró-núcleos, um masculino e outro feminino, levando à restauração do número diploide de cromossomos, ou seja, 46 cromossomos. Ao final desses eventos, forma-se o zigoto, e os 46 cromossomos que o formam se organizam em um fuso de clivagem.

O zigoto é composto por 23 cromossomos da mãe e 23 cromossomos do pai. Dessa forma, há uma nova combinação de cromossomos. Isso possibilita a variabilidade da espécie humana.
O zigoto é alvo de diversas clivagens, ou seja, divisões mitóticas repetidas, que se iniciam cerca de 30 horas após a fecundação e provocam um aumento do número de células rapidamente. Essas células são chamadas de blastômeros. A cada divisão formam-se blastômeros menores. Quando se formam nove blastômeros, os mesmos mudam sua forma e se unem, no processo de compactação, formando uma bola compacta de células. Essa proximidade das células permite a formação da massa celular interna.
Cerca de 3 dias após a fecundação forma-se a mórula, que, como já fora dito, é o primeiro estágio do desenvolvimento.
A mórula é composta por 12 a 32 blastômeros, sendo formada por uma camada interna que é a massa celular interna, e a camada interna é circundada por uma outra camada, que é a camada celular externa.
Esse processo de formação da mórula ocorre na tuba uterina.
No estágio de mórula, as células que a formam são células totipotentes. Mas, o que são células totipotentes? São células completamente indiferenciadas, capazes de gerar um organismo adulto. E essas células tem a capacidade de formar tanto um organismo adulto quanto tecidos extraembrionários, como a placenta.

BLÁSTULA
Quando a mórula chega ao útero, começa a surgir em seu interior uma cavidade, conhecida como cavidade blastocística. Essa alteração acontece devido à uma reorganização das células, que começam a ocupar a periferia, formando a cavidade. O interior dessa cavidade é preenchido por fluido, originado do útero, que penetra pela zona pelúcida. Essa cavidade é chamada também de blastocele. Nesse momento, o embrião deixa de ser chamado de mórula e passa a ser chamado de blástula.

A blástula, portanto, ao contrário da mórula, é oca, pois possui um espaço com líquido. À medida que o fluido aumenta na cavidade, ocorre a separação dos blastômeros em 2 tipos celulares: trofoblasto e embrioblasto.
Os trofoblastos correspondem a camada celular externa, a qual possui grande importância para formação da placenta.
Já os embrioblastos correspondem a camada celular interna e formarão o embrião propriamente dito. Posteriormente, o embrioblasto se diferencia em epiblasto e hipoblasto, formando o disco bilaminar.
No estágio de blástula as células da camada celular interna, os embrioblastos são células tronco pluripotentes, ou seja, são células capazes de gerar QUALQUER tecido do organismo. Porém, não são capazes de formar tecidos extraembrionários (pois essa capacidade é dos trofoblastos presentes na camada celular externa).
GÁSTRULA
Muitas alterações ocorrem, e chega-se, então, a uma nova etapa que sucede o estágio de blástula, essa etapa é a gastrulação que forma, portanto, a gástrula.
A gastrulação consiste no processo de formação das camadas germinativas. São três camadas germinativas e essas darão origem a todos os tecidos embrionários.
A gastrulação se inicia com o surgimento de uma linha conhecida como linha primitiva (originada por uma camada de células denominadas epiblastos, é uma diferenciação do embrioblasto que ocorre anteriormente ao processo de gastrulação). Ao se desenvolver, a linha primitiva gera algumas estruturas, como o nó primitivo (que corresponde a extremidade cranial da linha), o sulco primitivo (que é uma depressão na linha primitiva) e a fosseta primitiva (onde se encerra o sulco primitivo). Tanto o sulco quanto a fosseta primitiva são formados devido à invaginação de epiblastos.
Algumas células migram e formam o mesoderma, a camada intermediária.
Células do epiblasto deslocam as células hipoblásticas, e então o hipoblasto forma o endoderma, a camada mais interna.
E a camada de epiblasto forma o ectoderma, a camada mais superficial.
Assim, têm-se as três camadas: endoderma, mesoderma e ectoderma. Nesse estágio, o embrião é chamado de gástrula.
Nesse estágio, as células são células tronco multipotentes. Essas células tem a capacidade de se diferenciar em apenas alguns tipos de tecidos.
Cada uma das camadas é responsável pela formação de diversos tecidos. O endoderma forma, por exemplo, o Sistema Respiratório e Órgãos do Sistema Digestório. O mesoderma forma a derme, tecido conjuntivo e muscular, e o Sistema Circulatório e Reprodutor. E o ectoderma forma os epitélios, epiderme, cavidades, e o Sistema Nervoso.
Um evento importante que acontece nesse estágio e que será importante para o próximo, é a formação de uma estrutura a partir de células do mesoderma que formam então a notocorda, uma estrutura circular. A notocorda é importante para definir o eixo do embrião, estimular desenvolvimento do esqueleto axial, e de suma importância para o próximo estágio, de formação do Sistema Nervoso Central.
NÊURULA
No último estágio, eu disse que a notocorda seria muito importante para essa nova etapa.
A neurulação é o processo que dará origem ao sistema nervoso do embrião, que nesse estágio, é conhecido como nêurula. Esse processo é induzido/estimulado pela notocorda.
O sistema nervoso é derivado do ectoderma. O ectoderma localizado acima da notocorda sofre um espessamento, e, com isso, forma a placa neural. A partir daí, o ectoderma formador da placa neural passa a ser chamado de neuroectoderma, por ser uma “especialização” do ectoderma que dará origem ao sistema nervoso. A placa neural passa por várias alterações conformacionais, como formação de um sulco neural (uma depressão na placa), pregas neurais (que são proeminências da placa), até que essas pregas se unem e formam um tubo, conhecido como tubo neural. O tubo neural é o precursor do sistema nervoso central, formando encéfalo e medula espinhal. Quando esse tubo se forma, algumas células “ficam de fora” e formam, então, duas cristas neurais, que darão origem aos gânglios sensitivos dos nervos cranianos e espinhais. Além disso, contribuem na formação da retina, da glândula adrenal e etc.
Nesse momento, o embrião é chamado de nêurula.

Todos esses estágios acontecem até a quarta semana de desenvolvimento embrionário. Dá para acreditar que tantos acontecimentos importantíssimos acontecem em tão pouco tempo?!! Pois é, como valem quatro semanas para um embrião. São acontecimentos, que como você deve ter percebido, são cruciais para a vida. Por isso eu disse anteriormente, que anormalidades que possam vir a ocorrer nesses estágios iniciais podem inclusive inviabilizar a vida e formação do embrião, por serem fundamentais à vida!!
A vida de Mendel
Gregor Mendel foi um biólogo, botânico e monge que desenvolveu as bases da genética moderna. Os seus estudos o levaram a ser conhecido como o “pai da genética”.
Gregor Mendel nasceu em 20 de julho de 1822, na região da atual Áustria. Ele morreu em 6 de janeiro de 1884, por conta de uma doença renal.

Primeira lei de Mendel
A Primeira Lei de Mendel ou Lei da Segregação dos Fatores determina que cada característica é condicionada por dois fatores que se separam na formação dos gametas.
A segregação é consequência da localização dos genes nos cromossomos e do comportamento desses durante a formação dos gametas, através do processo de meiose.
O monge Gregor Mendel realizou seus estudos com objetivo de compreender como as diferentes características eram transmitidas de uma geração para outra.
Ausência de dominância
Herança intermediária
Co-dominância
Probabilidades em Genéticas
Fundamentos da probabilidade
- A probabilidade empírica de um evento é calculada contando-se o número de vezes que o evento ocorreu e dividindo esse valor pelo número total de vezes que o evento poderia ter ocorrido. Por exemplo, se o evento que você está buscando fosse o de uma ervilha com semente rugosa e você a viu 1, point, 850 vezes de um total de 7, point, 324 sementes examinadas, a probabilidade empírica de se ter uma ervilha de semente rugosa seria de 1, point, 850, slash, 7, point, 324, equals, 0, comma, 253, ou muito perto de 1 em 4 sementes.
- A probabilidade teórica de um evento, por sua vez, é calculada com base em informações sobre as regras e circunstâncias que produzem o evento. Esse tipo de probabilidade reflete o número de vezes que espera-se que um evento ocorra em relação ao número de vezes que ele poderia ocorrer. Por exemplo, se você tivesse uma planta de ervilha heterozigota para o gene da forma da semente (Rr) e a deixasse realizar autofecundação, você poderia usar as regras da probabilidade e seu conhecimento de genética para prever que 1 de cada 4 descendentes teria dois genes recessivos (rr) e seria rugosa, correspondendo a 0, comma, 25 (1, slash, 4) de probabilidade. Vamos discutir mais sobre como aplicar as regras da probabilidade neste caso.
A regra do produto
Uma regra de probabilidade que é muito útil na genética é a regra do produto, a qual afirma que a probabilidade de dois (ou mais) eventos independentes ocorrerem juntos pode ser calculada multiplicando-se as probabilidades individuais dos eventos. Por exemplo, se você rolar um dado de seis lados de uma vez, você tem uma chance de 1, slash, 6 de conseguir um seis. Se você rolar dois dados ao mesmo tempo, sua chance de conseguir duas vezes o seis é: (a probabilidade de um seis no dado 1) x (a probabilidade de um seis no dado 2) = left parenthesis, 1, slash, 6, right parenthesis, dot, left parenthesis, 1, slash, 6, right parenthesis, equals, 1, slash, 36.Em geral, você pode pensar na regra do produto como a regra do "e": se ambos evento X e evento Y devem acontecer para que ocorra um determinado resultado, e se X e Y são independentes um do outro (um não afeta a probabilidade do outro), então você pode usar a regra do produto para calcular a probabilidade do resultado multiplicando as probabilidades de X e Y.Podemos usar a regra do produto para prever a frequência de eventos de fertilização. Por exemplo, considere o cruzamento entre dois indivíduos heterozigotos (Aa). Quais são as chances de se ter um indivíduo aa na próxima geração? A única forma de se ter um indivíduo aa é se a mãe contribui com um gameta a e o pai contribui com um gameta a. Cada progenitor tem 1, slash, 2 das chances de fazer um gameta a. Logo, a chance de uma prole aa é: (probabilidade da mãe contribuir a) x (probabilidade do pai contribuir a) = left parenthesis, 1, slash, 2, right parenthesis, dot, left parenthesis, 1, slash, 2, right parenthesis, equals, 1, slash, 4.
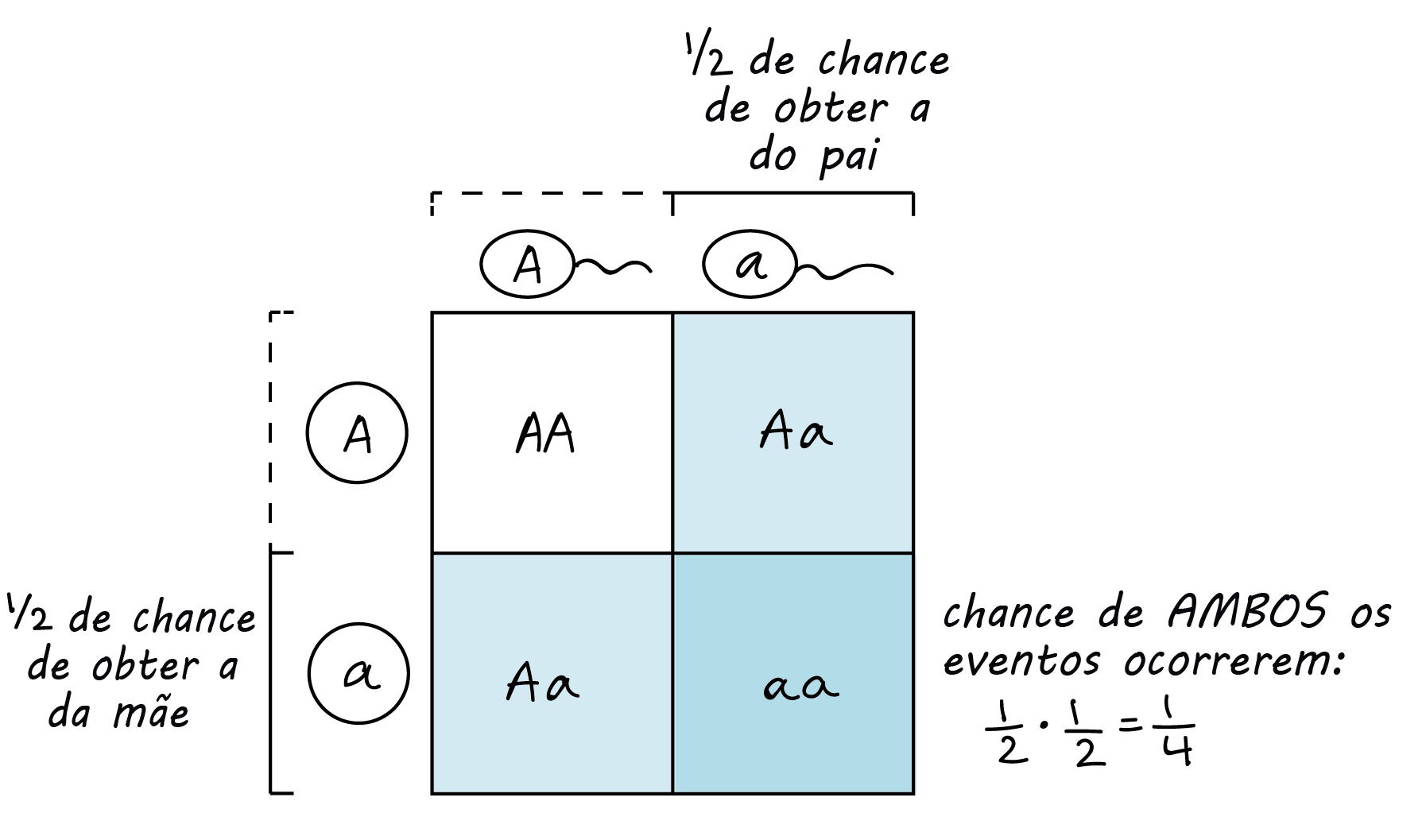
A regra do produto e a regra da adição
| Regra do produto | Regra da soma |
|---|---|
| Para eventos independentes X e Y, a probabilidade (P) de ambos ocorrerem (X e Y) é P, left parenthesis, X, right parenthesis, dot, P, left parenthesis, Y, right parenthesis. | Para eventos mutuamente excludentes X e Y, a probabilidade (P) de um ocorrer (X ou Y) é P, left parenthesis, X, right parenthesis, plus, P, left parenthesis, Y, right parenthesis. |

Comentários
Postar um comentário